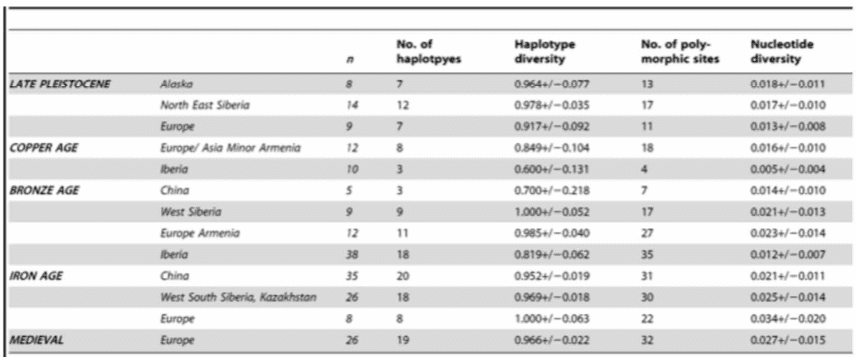
que aprender a cultivar porque es el que nos permite estudiar aquello que nos debe llamar la atención, nos dice que la cruza de individuos rayados con no rayados debería producirnos individuos menos rayados que el progenitor rayado, no más. Sin embargo, a la luz que arroja Bard, lo que importa es que si el caballo es de mayor envergadura que la cebra, específicamente si el embrión resultante es ligeramente más grande, la formación de rayas será más nutrida, lo que explica la interrogante.
Pero¿cómo es que un diseño aparentemente tan llamativo como el rayado de las cebras pudo fijarse tan marcadamente en tres especies equinas en África? Lo que es aún más impactante, cómo es que este diseño extravagante no es visto en ninguna otra parte del mundo?, ¿por qué sólo en África? Debido a que sólo de manera ocasional algunos caballos en otras partes del mundo desarrollan rayas en las patas que no guardan ninguna relación con las rayas de las cebras, podemos estar seguros que el diseño rayado no es primitivo, sino una invención africana.
Josef H. Reichhof en La Aparición del Hombre (Crítica 1996) aborda este problema en dos capítulos. La primera pregunta del párrafo anterior recalca lo llamativo del diseño rayado de alto contraste blanco y negro. Los animales, sin importar su lugar en la cadena alimenticia, tienden a beneficiarse si no llaman la atención. Claro, existen excepciones, como cuando es vital atraer al sexo opuesto, o al polinizador (que normalmente se guía por patrones ultravioletas y no visibles), o como cuando una mariposa anuncia su espantoso sabor con llamativos colores, si algún depredador comete el error de comerse a un representante de esa especie seguramente jamás lo volverá a hacer, pero las cebras tienen fama de ser exquisitas, por lo que su fácil identificación generaría el efecto contrario. De hecho, las cebras son fácilmente identificadas por los leones, que las acechan desproporcionadamente. Sin embargo, sus poblaciones no tienden a sufrir por la desventaja que hacia sus depredadores les plantea su estilo único.

Leona cazando una cebra.





 21.52.19")
 Sabemos que los caballos, famosos en México porque aparentemente los indígenas al verlos por primera vez en batalla no sabían si estaban peleando contra centauros (vamos, pensaban que eran una sola criatura y no un hombre anarmadura montado en una bestia) o deidades de algún tipo (ver, e.g., La Conquista de México, de Hugh Thomas [2000], editorial Planeta pg. 205). Lo que muchos mexicanos (o en todo caso americanos) no saben –y deberían- es que el caballo se originó en América. Efectivamente, el caballo nació en los parajes norteamericanos aproximadamente unos 2 millones de años atrás, aunque su linaje normalmente se traza 50 millones de años en el pasado, concretamente en la humilde (y casi canina) figura de
Sabemos que los caballos, famosos en México porque aparentemente los indígenas al verlos por primera vez en batalla no sabían si estaban peleando contra centauros (vamos, pensaban que eran una sola criatura y no un hombre anarmadura montado en una bestia) o deidades de algún tipo (ver, e.g., La Conquista de México, de Hugh Thomas [2000], editorial Planeta pg. 205). Lo que muchos mexicanos (o en todo caso americanos) no saben –y deberían- es que el caballo se originó en América. Efectivamente, el caballo nació en los parajes norteamericanos aproximadamente unos 2 millones de años atrás, aunque su linaje normalmente se traza 50 millones de años en el pasado, concretamente en la humilde (y casi canina) figura de